
L'EVOLUZIONE DEI MOLLUSCHI BIVALVI [parte 01]
Pubblichiamo, con grande piacere, la prima parte della tesina di Piva Lavinia, dedicata all'evoluzione dei molluschi bivalvi
01.RIFERIMENTI FILOSOFICI DELLA TEORIA DELL'EVOLUZIONE DI DARWIN
L'evoluzione biologica può essere descritta come una serie di cambiamenti che portarono delle specie esistenti in passato a originare quelle attualmente presenti sulla terra.
Nella seconda metà dell'800 si affermò l'idea di un'evoluzione della vita, dopo che Charles Darwin spiegò in base a dettagliate osservazioni, come le specie cambiano nel tempo.
Egli utilizzò dati derivanti dallo studio dei fossili e delle anatomie rappresentate, come prove a favore della sua tesi evolutiva, e si rese conto che le specie attuali non derivano le une dalle altre, ma possono avere un antenato comune.
Durante la sua vita, Darwin intraprese un lungo viaggio durato 5 anni, dove poté studiare la biologia e la geologia di regioni distanti nel globo.
In particolare alle Galàpagos osservò la presenza di specie animali differenti tra loro, ma simili per alcune caratteristiche, come ad esempio le tartarughe terrestri giganti, la cui forma del carapace differiva da una zona all’altra dell’isola, o come un gruppo di volatili che si distinguevano tra loro per la forma del becco.
Fu qui che Darwin iniziò ad avere i primi dubbi sulla teoria del fissismo, secondo cui le varie specie possiedono caratteri che le definiscono fin dall'origine (a partire dalla creazione divina), espressa da alcuni uomini di chiesa del tempo.
Egli, non accettando questa idea, ritenne che ci fosse una relazione instauratasi tra ambiente e organismi.
Tutti gli individui appartenenti a una particolare specie hanno molteplici caratteri comuni, ma nonostante ciò, essi sono differenti gli uni dagli altri.
Per ogni specie esiste una variabilità individuale casuale fondamentale per l'evoluzione; in molti organismi la progenie non è uguale né a sé stessa, né a chi l'ha generata. Darwin ipotizzò che il tasso di sopravvivenza e di riproduzione degli individui dipendesse da quelle leggere differenze. [Nella sua opera fondamentale "L'origine delle specie" (1859) scritta dopo il suo viaggio attorno al mondo, egli realizzò una vera e propria rivoluzione scientifica. Il libro, appena pubblicato, scatenò accesi dibattiti e contemporaneamente entusiasmo da parte di alcuni lettori].
Darwin inoltre trovò insoddisfacente la teoria evoluzionista formulata da Jean-Baptiste de Lamarck, zoologo francese che sostenne che le specie subiscono un'evoluzione grazie agli stimoli dell’ambiente, perché non aveva una base scientifica argomentata e verificata. Per il filosofo la scienza doveva fornire spiegazioni accettabili delle teorie.
La variazione delle condizioni ambientali e l’accrescimento del numero degli individui di una stessa specie pongono agli organismi problemi di adattamento che li spingono a una lotta per l’esistenza: chi si dimostrava più adatto alle nuove condizioni sopravviveva.
Attraverso le analisi del materiale raccolto dalle sue osservazioni, Darwin formulò la legge della "selezione naturale", che fu la risposta all’interrogativo sulle modalità dell'evoluzione biologica.
La teoria della "selezione naturale" di Darwin è riassunta nei seguenti punti:
- alcuni organismi producono variazioni nella loro struttura biologica che li rendono più adatti a determinate condizioni ambientali;
- tali organismi nella lotta per l’esistenza sono quelli che sopravvivono, mentre gli altri si estinguono;
- in essi si stabilizzano nuovi caratteri che vengono acquisiti irreversibilmente, trasmettendosi ereditariamente ai discendenti;
- i caratteri ereditari possono rappresentare una vera e propria mutazione di una determinata specie;
- l'evoluzione delle specie non è immediata ma graduale, e implica la nascita di molte specie intermedie di cui si ha testimonianza attraverso reperti fossili.
In conclusione per Darwin le variazioni insorgono in modo casuale e, successivamente, è l'attività dell'ambiente che opera una selezione favorendo i gruppi con mutazione che risultano più utili, portando alla scomparsa dei gruppi che ne sono privi.
02.L'EVOLUZIONE DEI MOLLUSCHI
Prima di entrare nel merito della evoluzione dei bivalvi è necessario fornire una breve descrizione del phylum dei molluschi, del quale i bivalvi fanno parte e della loro filogenesi.
Ho preso spunto da varie fonti ed in particolar modo ho effettuato una libera traduzione del testo Phylogeny and evolution of bivalve molluscs - Elizabeth Gosling.
Il phylum Mollusca è uno dei gruppi più grandi e più diversificati del regno animale, con almeno 50.000 specie descritte e probabilmente ben 200.000 specie viventi, la maggior parte delle quali sono marine.
Il phylum ha un considerevole primato di fossili, andando indietro sino al Cambriano, cioè sino a circa 540 milioni di anni fa (Ponder & Lindberg, 2008).
I molluschi sono animali dal corpo molle, e la maggior parte di loro ha un guscio protettivo duro, chiamato conchiglia, all'interno della quale vi è una grande struttura di tessuto chiamata mantello. Il mantello racchiude gli organi interni dell'animale. Un'altra caratteristica del phylum è un grande piede muscolare che viene generalmente usato per la locomozione. Sebbene la maggior parte dei molluschi condividano queste caratteristiche di base, il phylum è caratterizzato da una grande diversità di forme e comportamenti. Si va dalle patelle aggrappate alle rocce, alle lumache che strisciano, a bivalvi che si ancorano, scavando nel substrato o forano le rocce, ai cefalopodi che sfrecciano nell'acqua o si nascondono, mimetizzandosi sul fondale.
Essi occupano tutti gli habitat: gli abissi del mare, le barriere coralline, le distese fangose, foreste, fiumi, laghi e vivono anche sotto terra. Essi possono anche essere nascosti come parassiti all'interno di altri animali, si nutrono di ogni possibile alimento e sono di dimensioni variabili, dai calamari giganti alle vongole, a piccole lumache lunghe un millimetro.
03.FILOGENESI DEI MOLLUSCHI
Un sistema di classificazione è basato sulla storia evolutiva dei gruppi di organismi, piuttosto che sulle caratteristiche puramente condivise. Sono conosciute otto classi di molluschi (Figura 1.1), per lo più basate sulla tassonomia (filogenetica) e sull'analisi dei caratteri morfologici di taxa viventi e fossili.
La classe Aplacophora conterrebbe due sottoclassi: Solenogastres o Neomeniomorpha (~ 250 specie) e Caudofoveata o Chaetodermomorpha (~ 150 specie), si tratta di animali a forma di verme delle acque profonde privi di una conchiglia, ma coperti da una cuticola e da spicole di aragonite. La classe Polyplacophora (~ 100 specie), spesso definiti come chitoni, vivono su substrati duri sulle coste rocciose, e sono caratterizzati da otto piastre dorsali.
Le classi Aplacophora e Polyplacophora sono raggruppati nel subphylum Aculifera, che è considerato monofiletico, cioè tutti i taxa in questo gruppo condividono un antenato comune (Sigwart & Sutton 2007).
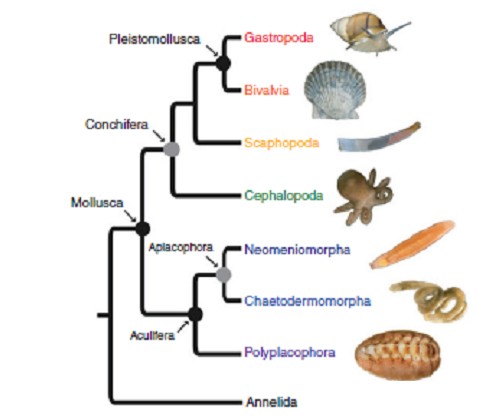
Fig 1.1. Albero filogenetico dei molluschi, creato sulla base dei dati del trascrittoma e del genoma di tutte le principali classi, ad eccezione dei Monoplacophora; i cerchi neri rappresentano i nodi con il supporto del metodo statistico bootstrap (bs) = 100 e probabilità a posteriori (pp) = 1,00; i cerchi grigi rappresentano i nodi con bs = 100 e ppa ≥0.98.
Il Bootstrap viene utilizzato per valutare la stabilità dei raggruppamenti in un albero filogenetico, con successive misure della probabilità (la probabilità che un evento si verifichi, dato che un evento simile si è già verificato).
Ci sono solo circa 30 specie della classe Monoplacophora (non mostrate in figura 1.1) e tutte vivono in acque profonde. Sono piccole, simili a patelle e con un unico guscio di protezione. La classe Scaphopoda (~ 600 specie), o conchiglie a zanna, compende molte specie che vivono nel fango e nei sedimenti marini.
La classe Scaphopoda (~ 600 specie), comunemente nota come conchiglie a zanna, a causa della loro guscio conico e un po' curvato, le specie relative a questa classe vivono nel fango e nei sedimenti marini.
La classe Gastropoda è la più numerosa (> 100.000 specie) ed è la più diversificata; comprende chiocciole con guscio a spirale arrotolata, patelle piatte, nudibranchi senza guscio, chiocciole e lumache terrestri.
La classe Bivalvia, con circa 9.200 specie (Huber, 2010), include gli animali chiusi in due valve, quali cozze, ostriche, capesante e vongole.
Polpi, calamari, seppie e Nautilus appartengono alla classe Cephalopoda. Ci sono circa 1.000 specie in questa classe e rappresentano la classe più organizzata e specializzata di tutti i molluschi. Queste quattro classi sono raggruppate nei Conchifera, che è considerato come un gruppo monofiletico.
I mollusco ancestrale ipotetico sia un evoluto verme piatto, oppure un anellide dalla struttura semplice. Si pensa che sia stato un animale di piccole dimensioni (1-3 mm), vissuto nei mari poco profondi del Precambriano, e che strisciando sopra il substrato su un grande piede, raschiava le alghe dalle rocce con il suo apparato boccale specializzato. Nella parte posteriore dell'animale vi doveva essere una coppia di ctenidia (branchie) filamentosi e ciliati, che funzionavano solo come organi respiratori.
Se una tale creatura sia realmente esistita è comunque un fatto controverso; fino a tempi relativamente recenti i dati morfologici erano l'unica fonte utilizzata per dedurre relazioni filogenetiche all'interno dei molluschi.
Nel 1980 l'applicazione dei metodi molecolari hanno rappresentato un avanzamento potenzialmente importante per chiarire le relazioni di questo importante phylum.
La maggior parte delle analisi si sono concentrate su singoli geni nucleari, tuttavia le fondamentali domande sull'evoluzione dei molluschi sono rimaste in gran parte senza risposta da parte dei metodi morfologici e da dati molecolari, che spesso hanno dato risultati non congruenti.
Ad esempio non è chiaro se ì vermiformi Aplacophora si siano diversificati prima dei Conchifera o abbiano perso i loro gusci, oppure se i Polyplacophora siano un gruppo affine ai Conchifera o ai Monoplacophora e quali siano le interrelazioni dei gruppi di Conchifera.
Recentemente i ricercatori hanno adottato un approccio multigene nel tentativo di rispondere a queste domande. Questo approccio utilizza sequenze derivate da dati sui genomi, che consentono di codificare numerosi geni codificanti proteine, che possono essere identificati e impiegati nella ricostruzione della filogenesi.
Questo approccio utilizza sequenze derivate da dati sui genomi, che consentono di sequenziare numerosi geni codificanti proteine o producenti RNA non codificante, che possono essere identificati e impiegati nella ricostruzione della filogenesi. Questi studi hanno portato a sostenere l’esistenza di un rapporto di stretta affinità tra gasteropodi e bivalvi, proponendo il nome di Pleistomollusca per questo raggruppamento, che contiene più del 95% di tutte le specie di molluschi (Figura 1.1).
Il supporto per questo raggruppamento (clade) si basa principalmente su analisi molecolari, anche se sono state proposte alcune sinapomorfie (caratteri derivati condivisi) morfologiche, quali: muscoli larvali retrattili, membrana muscolare ad anello, e forse la perdita della radichetta ciliare anteriore nelle loro ciglia locomotorie.
La classe Scaphopoda sarebbe affine al clade Pleistomollusca, anche se le conferme sarebbero meno solide.
I cefalopodi sarebbero quindi una classe affine a tutti gli altri Conchifera o Placophora. (Smith et al. 2011), utilizzando un approccio simile, questa volta comprendendo anche i Monoplacophora, hanno trovato un forte sostegno per la definizione dei due principali cladi, ritenendo però che tra i molluschi con conchiglia, i Monoplacophora non sarebbero un gruppo affine a tutti gli altri Conchifera, come era stato suggerito dalla maggior parte degli autori, ma sarebbero invece un gruppo affine ai cefalopodi. Questi autori hanno anche trovato supporto per la definizione di un clade composto da Bivalvia, Scaphopoda e Gastropoda, con gli ultimi due gruppi caratterizzati da maggiore affinità.
A dimostrazione della estrema complessità nel definire la filogenesi dei molluschi, (Vinther et al., 2012), utilizzando sette geni nucleari, hanno trovato un supporto alla tesi secondo la quale i cefalopodi sarebbero invece un gruppo affine agli Aculifera.
FINE PRIMA PARTE
AGGIUNGI UN COMMENTO [Se non desideri visualizzare il tuo commento sul tuo profilo Facebook, ricordati di togliere la spunta della ceckbox]
ARTICOLI CORRELATI
- L'EVOLUZIONE DEI MOLLUSCHI BIVALVI Parte 2
- L'EVOLUZIONE DEI MOLLUSCHI BIVALVI Parte 3
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
