
LE ALOFITE
In base alla disponibilità di acqua le piante sono classificate in idrofite (vivono in ambienti ove l’acqua è sempre disponibile), mesofite (con disponibilità irregolare di acqua) e xerofite (che vivono in ambienti con scarsa disponibilità di acqua). Queste ultime sono rappresentate dalle piante che vivono in ambienti estremamente aridi come i deserti. Questi ambienti sono rappresentati da condizioni mutevoli e con potenziali idrici estremamente variabili da luogo a luogo. Per ovviare a questi problemi, le piante possono evitare potenziali idrici estremamente negativi, come fanno per esempio le palme delle oasi nei deserti, che attingono alla falda attraverso un apparato radicale molto sviluppato; altre piante sfuggono alla siccità (piante effimere dei deserti) e, nella stagione secca, si trovano solo come semi disidratati; le succulente a metabolismo CAM accumulano acqua che poi utilizzano in modo estremamente parsimonioso, riducendo la traspirazione grazie alla loro cuticola molto spessa e chiudendo gli stomi durante il giorno.
Questo delle succulente è un altro modo per evitare la siccità; le piante economizzatrici di acqua non succulente hanno sviluppato altre strategie per limitare la perdita di acqua, per esempio hanno lamine fogliari piccole, in modo tale da aumentare la perdita di calore per convenzione e abbassare la temperatura e, di conseguenza, ridurre la traspirazione. Hanno anche stomi infossati e peli sulla superficie delle foglie che possono essere perse durante la stagione secca.
Altre piante infine resistono alla siccità, e queste sono estremamente interessanti per i botanici tanto che sono chiamate xerofite vere o euxerofite, poiché a differenza delle altre non sfuggono e non evitano la siccità, bensì resistono ad essa nel vero senso del termine. Anche queste piante presentano stomi infossati o foglie piccole, ma hanno evoluto adattamenti fisiologici eccezionali; alcune di esse possono seccarsi, reidratarsi e divenire di nuovo metabolicamente attive, anche dopo molto tempo, e sono conosciute con il nome di poichiloidriche. Alcune piante desertiche sono eteroblastiche (vedere paragrafo eteroblastia), producono cioè semi morfologicamente diversi, in modo tale da poter germinare all’interno di uno spettro di condizioni ambientali estremamente variabili.
Naturalmente i soluti presenti nei terreni influenzano il potenziale idrico e possono essere molto tossici per le piante, così in base alla capacità di tollerare o meno i soluti, le piante sono suddivise in glicofite (sensibili a concentrazioni più o meno elevate di sali) e alofite (che sopravvivono anche a elevate concentrazioni di sali). Anche per le alofite è possibile fare una distinzione tra a. eurialine e a. stenoaline; le prime tollerano variazioni ampie della salinità, le seconde crescono meglio su substrati a salinità costante.
Le alofite, termine che deriva dal greco halo = sale e phyte = pianta (in inglese halophytes plants oppure salt water plants), sono quindi in grado di portare a termine il proprio ciclo di crescita in condizioni ambientali caratterizzate da elevata salinità, per la presenza di sodio, cloro, potassio e carbonati, derivati questi ultimi dai gusci dei molluschi spiaggiati. Esse affondano le loro radici in terreni quasi sempre riducenti, che trattengono poco o nulla l’acqua meteorica che ricevono, resistono con adattamenti che esamineremo, a condizioni climatiche estreme; debbono acquisire acqua da un suolo con potenziale osmotico negativo; infine, debbono smaltire ioni come sodio, carbonato e cloruro che possono essere tossici quando presenti in eccesso.
Senza addentrarci in aspetti troppo nozionistici, ricordiamo che l'ambiente delle spiagge è caratterizzato da elevata salinità (alte concentrazione di Na+ e minore disponibilità di Ca2+ e Mg2+ al suolo). La concentrazione salina delle soluzioni presenti in un substrato quale la spiaggia, può arrivare anche al 10% (la saturazione si ha attorno al 38%). Il pH in genere è inferiore a 8.5.
Nei testi anglosassoni, spesso le aree costiere sono classificate come umide (wet saline area), che si contrappongono alle zone saline secche dei deserti (dry saline area). L’umidità delle spiagge è garantita sia dal moto ondoso e dagli aerosol, ma anche a pochi cm di profondità, dalla risalita del cuneo salino, tanto maggiore quanto più si riduce l’apporto idrico delle falde di acqua dolce o la portata dei fiumi. C'è, infine, un altro aspetto da considerare, il potenziale osmotico dell’acqua che è instabile e può innalzarsi bruscamente; per esempio, dopo intense precipitazioni che addolciscono l’acqua necessaria alla crescita delle piante, oppure dopo intensa evaporazione, si può avere un brusco innalzamento della concentrazione salina. Le angiosperme alofite che esamineremo sono quelle adattate agli ambienti variabili delle coste sabbiose, ghiaiose e rocciose, mentre sono escluse le angiosperme marine vere e proprie (alghe come Posidonia, Cymodocea e Zoostera).
CLASSIFICAZIONE
Sulla base della resistenza e della tolleranza al sodio, le alofite sono classificate come segue (1):
- obbligate, o vere alofite o eualofite, resistenti a condizioni estreme di salinità (> 1.5 %);
- transitorie, che caratterizzano gli ambienti di transizione, ossia gli ambienti intermedi tra suoli salini e suoli non salini;
- facoltative, che crescono in suoli non salini, ma che tollerano abbastanza bene un certo grado di salinità, in genere non superiore a 0.5 %.
Altra classificazione basata sulla risposta allo stress salino considera le eualofite (o mioalofite), come quelle specie capaci di crescere su terreni con salinità superiore a 720 mM. Le eualofite vivono in terreni saturi di sali, ma possono benissimo crescere anche in terreni tipici delle glicofite. Secondo alcuni ricercatori, nessuna alofita è veramente obbligata, ma sono rare in terreni non salati perchè non sono in grado di competere efficacemente con le glicofite.
Per concludere, citiamo una ulteriore suddivisione delle alofite, per cui si hanno da una parte le idroalofite, che vivono perennemente su suoli umidi come le mangrovie o la nostra Praghmites e, dall’altra, le xeroalofite, che vivono sempre su suoli salini che possono divenire aridi in certi periodi dell'anno. Si tratta di un gruppo di piante confinate ai deserti.
La figura seguente mostra l’andamento della crescita in funzione della concentrazione espressa in M di NaCl per quattro gruppi di piante:
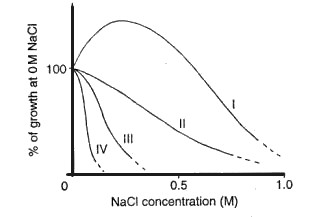
Il gruppo indicato con I comprende le alofite estreme o eualofite, capaci di completare il ciclo di crescita in ambienti estremamente salati, comprende generi come Suaeda, Salicornia e Atriplex. Il gruppo II comprende alofite transitorie come le dicotiledoni Aster tripolium e Cochlearia anglica e le monocotiledoni come Puccinellia maritima. Il gruppo III include le alofite facoltative (o se vogliamo le glicofite alotolleranti o pseudoalofite, tendono ad accumulare sali a livello del parenchima xilematico). Infine il gruppo IV comprende le normali glicofite. Figura tratta da: Dunes Ecology.
ADATTAMENTI EVOLUTIVI NELLE ALOFITE
Tra i fattori edafici che maggiormente influenzano la distribuzione delle specie vegetali, vi sono, oltre al contenuto di acqua, anche il contenuto di sali; la tossicità ionica incide notevolmente a livello di germinazione, crescita e sviluppo delle piante. Gli effetti di una concentrazione elevata di soluti ionici sono percepiti dalle piante allo stesso modo di uno stress idrico, e si parla in questo caso di aridità fisiologica.
Gli effetti tossici si manifestano su tre principali livelli, e cioè (I) a livello cellulare con alterazione dei sistemi di membrane dei mitocondri e dei cloroplasti, nonché con alterazione dei sistemi enzimatici, (II) a livello dei tessuti, (III) a livello dell’intera struttura della pianta, per alterazione dell’ingresso, del trasporto e della distribuzione dei sali nutritivi e conseguente riduzione della crescita dell’apparato fogliare, radicale, dei fiori e dei germogli.
I suoli ricchi di sale influenzano naturalmente anche l’apporto idrico, poiché tali suoli presentano un potenziale idrico estremamente negativo. Sodio e cloro sono i soluti ionici che penetrano eccessivamente a discapito di calcio e potassio, e causano una serie di effetti conosciuti con il termine di salt stress. Naturalmente, questi effetti sono riconducibili alle glicofite, mentre le alofite hanno evoluto una serie di meccanismi di protezione atti ad evitare o limitare le conseguenze dello stress salino.
A livello ecofisiologico è stato introdotto negli anni '80 il termine di osmoregolazione (osmo-regulation o osmotic adjustment), per indicare la capacità delle alofite di utilizzare soluti ionici dell’ambiente esterno o composti organici sintetizzati al fine di mantenere il giusto potenziale idrico, che deve essere minore rispetto al compartimento extracellulare e a quello del suolo. Per salt includers si intendono i composti inorganici osmoticamente attivi quali sodio, cloro, potassio e calcio, accumulati selettivamente all’interno delle cellule. Tali alofite, sono dette regolatrici di sali. Il contenuto di sodio, cloro e potassio nelle alofite, è compreso in un range abbastanza variabile e in generale il rapporto Na:K è basso nelle monocotiledoni e più elevato nelle dicotiledoni (Flowers et al. 1986). Inoltre la concentrazione dei soluti ionici è bassa nel citosol mentre risulta elevata all’interno dei vacuoli. Per esempio, sempre secondo il lavoro di Flower et al. del 1986, in Suaeda maritima, il contenuto di sodio, potassio e cloro è così distribuito (espresso in mM):
Na |
K |
Cl |
|
esterno |
340 |
7 |
340 |
citoplasma |
166 |
27 |
86 |
vacuolo |
494 |
20 |
352 |
Il basso contenuto citoplasmatico di soluti inorganici nel citoplasma, tipico di molte alofite, non spiega da solo il valore del potenziale idrico dell’acqua. In realtà, sono stati trovati da tempo diversi composti organici che contribuiscono al mantenimento del potenziale osmotico del citosol, tra questi, composti quaternari dell’ammonio come la glicin-betaina, composti terziari dello zolfo, zuccheri, mannitolo, sorbitolo, galattosil-glicerolo, vari acidi organici e poliammine. Queste molecole sono comunemente indicate come osmoprotettori o soluti compatibili (compatible solute). Tra queste, la prolina sembrerebbe svolgere un ruolo duplice, e cioè quello di soluto compatibile e come fonte di azoto o di amminogruppi. Si ritiene che il principale ruolo di queste molecole sia tuttavia quello di ridurre le specie reattive dell’ossigeno (ROS, reactive oxygen species), prodotte e attive maggiormente a livello de cloroplasti.
La sintesi dei sistemi enzimatici sensibili al cloruro di sodio (PEP carbossilasi, malato deidrogenasi, nitrato reduttasi, glucoso-6-fosfato deidrogenasi, perossidasi, ATPasi ecc..), sembra essere in relazione alla concentrazione salina. Sempre in Suaeda maritima è stata dimostrata in vitro, già verso la fine degli anni ‘70, una diminuzione di alcuni sistemi enzimatici con l’aumento della concentrazione di NaCl. Tuttavia rimane da verificare se il comportamento in vitro è paragonabile al comportamento in vivo. L’abilità delle piante alotolleranti sta nell’avere evoluto meccanismi di controllo della concentrazione dei soluti, attraverso (I) la prevenzione dell’accumulo di sodio, (II) la riduzione della concentrazione di sodio, (III) attraverso la sua compartimentazione ed espulsione.
L’ingresso nei vacuoli di NaCl avviene attraverso un sistema di antiporto H+/Na+ ATPasi-dipendente. Nella alofita Atriplex nummularia, è stato dimostrato in laboratorio che dopo l’aggiunta a intervalli regolari di 400 mM di NaCl, la sintesi di mRNA del sistema H+/Na+ ATPasi-dipendente aumenta regolarmente. Ciò dimostrerebbe la presenza di elementi di trascrizione e/o di fattori di trascrizione unici delle alofite, per lo meno nella specie oggetto di studio (4). Un’altra interessante caratteristica delle alofite, riguarda la capacità che esse hanno di discriminare tra sodio e cloro, altre ancora hanno adottato la strategia di mantenere inattivi i canali ionici per la maggior parte del tempo, oppure è limitato il numero totale per unità di superficie dei canali stessi. Nella specie Suaeda maritima, i canali SV (slow vacular ione channel) voltaggio-dipendenti hanno una probabilità di apertura tanto minore quanto maggiore è il decremento negativo del potenziale del tonoplasto. Non risulta invece chiaro se l’accumulo di acido abscissico che è stato riscontrato nelle glicofite sottoposte a stress salino, avvenga anche nelle alofite.
Tra gli adattamenti tissutali, il più noto è quello della succulenza che porta all’accumulo di grandi quantità di liquidi e conseguente rigonfiamento delle foglie (Suaeda) o dei fusti (Salicornia), e aumento del rapporto superficie-volume. L’accumulo determina una diluizione dei soluti tossici, semplicemente perchè si accresce la quantità di solvente e rimane pressoché costante quella dei soluti ionici. Altre piante adattano la velocità di accrescimento all’accumulo di sali, in tal modo rimane costante la concentrazione ionica. Queste alofite sono dette regolatrici di sali. Strutture specializzate nell’eliminazione o nell’accumulo dei sali in eccesso sono rispettivamente le salt gland e le salt hirs. Si tratta di strutture dette salt exclusion structure, proprie delle secretoalofite. Le esoghiandole saline o salt glands, sono state descritte per la prima volta nella famiglia delle Tamaricaceae (Marloth, 1887) e nelle Plumbaginaceae (Volkens, 1884); si attivano solo in condizioni di elevata salinità e sono presenti anche in piante non alofite. Si trovano in almeno 11 famiglie di dicotiledoni e una di monocotiledini (Graminaceae). Sono presenti in Limonium, Spartina, Crypsis, Digitaria, Eranthius. Sono disposte quasi sempre entro depressioni dell’epidermide nelle parti aeree della pianta. Le endoghiandole saline o bladder cells o bladder hairs scoperte solo nel 1959, svolgono la stessa funzione delle esoghiandole. Sono presenti in molti generi della famiglia Chenopodiaceae (Atriplex, Halimione, Chenopodium) e si presentano come peli (tricomi) a livello delle parte aeree dell’epidermide che accumulano sali. Dai tessuti fogliari il sale viene trasportato attraverso la piccola cellula del peduncolo alla cellula vescicolare rigonfia. Con il tempo aumenta la concentrazione di sale fino a determinare l’esplosione della cellula stessa, che cade liberando il suo contenuto.
Un meccanismo simile, quello cioè di eliminare sali in eccesso dalla superficie fogliare, è utilizzato da alcune alofite allo scopo di assorbire igroscopicamente acqua dall’umidità dell’aria (es. Nolan mollis, vedere lavoro di Mooney et al. 1980). Quindi in tal caso l’acqua è assorbita direttamente dalle foglie, o indirettamente dalle radici che utilizzano la soluzione salina sgocciolata a terra, ambedue le vie richiedono energia metabolica e sono comunque poco conosciute. Queste piante al termine del ciclo vitale decomponendosi arricchiscono i terreni circostanti di sali influenzando fortemente l’ambiente in cui crescono. Il lavoro di Fireman e Hayward del 1952, dimostrò che la specie Sarcobatus vermiculatos che cresce nel deserto dello Utha, preleva sali in profondità e li rilascia in superficie attraverso la caduta delle foglie, saturando di sali il terreno circostante. Altre piante accumulano invece ioni in tessuti senescenti, per esempio Salicornia accumula ioni nei germogli e nelle foglie.
Nei terreni poveri di calcio l’assorbimento di ioni sodio entra in competizione con l’assorbimento di ioni potassio attraverso un meccanismo di bassa affinità. Solo se la quantità di calcio è sufficiente, lo ione potassio è assorbito adeguatamente attraverso un meccanismo di alta affinità, che trasporta prevalentemente potassio in luogo dello ione sodio (Le Haye & Epstein 1979). Il meccanismo del calcio che limita gli effetti dannosi del sodio è stato studiato ampiamente, e probabilmente sono coinvolti meccanismi di trasporto attraverso le membrane.
Un’altra caratteristica morfologica di alcune alofite riguarda la dislocazione degli stomi, posti spesso in una depressione in modo tale da poter sfruttare l’elevata pressione parziale di vapore dell’aria che vi rimane intrappolata. Altre piante, quelle a metabolismo CAM, chiudono gli stomi di giorno per limitare la traspirazione, così acquisiscono la CO2 di notte legandola a particolari acidi organici e accumulandola nei vacuoli, effettuando la fotosintesi di giorno a stomi chiusi. Ma la particolarità più interessante, riguarda la capacità che le cellule di guardia proprie di alcune alofite. Tali cellule sono adattate a elevate concentrazioni saline e utilizzano il sodio in luogo del potassio per il mantenimento del turgore cellulare, così come accade in Suaeda maritima. In altri casi, il ruolo del potassio è limitato oppure risulta il catione dominante come in Aster tripolium. In tal caso le cellule di guardia disporrebbero di un meccanismo atto a limitare l’ingresso di sodio.
Nel complesso le alofite hanno sviluppato adattamenti di varia natura che possiamo così riassumere:
- regolazione e/o compartimentazione dei soluti ionici;
- adattamenti osmotici e produzione di osmoliti;
- adattamenti enzimatici;
- adattamenti degli stomi;
- adattamenti della germinazione;
- adattamenti tissutali e morfologici di varia natura, come impermeabilizzazione delle radici; succulenza; riduzione delle superfici fogliari; aumento dello spessore delle foglie; alterazione del colore delle foglie;; riduzione del numero e della grandezza delle foglie; necrosi marginale; sviluppo di ghiandole specializzate per l’escrezione e l’accumulo dei soluti in eccesso; morfologia della pianta adattata alle condizioni locali (large tussocks a ciuffo espanso, tuffed growth crescita a ciuffo, large branched forms a ramificazione espansa, small branched forms a ramificazione poco espansa, large erect forms forma eretta, small erect forms forma eretta piccola, large rosette forma a rosetta espansa, small rosette forma a rosetta contratta, prostrate forms forma prostrata. Si tratta di termini che spesso si incontrano in letteratura). Vedere tabella sottostante per spiegazioni;
- adattamenti fisiologici, come riduzione dell’assimilazione della anidride carbonica, passaggio del metabolismo da C3 a CAM (es Mesembryanthemum crystallinum); incremento a bassa salinità dell’efficienza del sistema radicale attraverso lo sviluppo di micorrizie.
MORFOLOGIA |
CARATTERISATICHE |
||
large tussocks |
|
||
tuffed growth |
|
||
large branched form |
tallo oltre 25 cm. Molto ramificata. Es Sonchus asper. |
||
small branched form |
Come la precedente ma su scala ridotta. Es Salsola kali. |
||
large erect forms |
|
||
small erect forms |
Come la precedente ma su scala ridotta e tallo infeiore a 25 cm. Es. Linum catharticum. |
||
large rosette |
|
||
small rosette |
Come la precedente su scala ridotta. Inflorescenze senza o con pochissime foglie. Es Taraxacum officinale. |
||
prostrate forms |
Piante prostrate, che si propagano cioè da un centro verso l’esterno. Es. Sedum acre. |
GERMINAZIONE e ETEROBLASTIA
I semi delle alofite rispondono agli stress salini in modo simile a quelli delle glicofite (il massimo della germinazione si ha per i due gruppi di piante in acqua distillata), tuttavia essi sono capaci di tollerare elevate concentrazioni saline per tempi anche molto lunghi, e dare avvio alla germinazione in condizioni favorevoli. La soglia di salinità oltre la quale si ha l’inizio del processo di germinazione, dipende da specie a specie. Alle medie latitudini lo stress salino è massimo durante i periodi estivi, quando l’intensa evaporazione lascia al suolo una coltre di sale ben evidente, sopratutto lungo i canali di marea lagunari e in generale, nelle aree allagate degli ambienti costieri. Questo evidenzia anche la notevole capacità dei semi delle alofite di rimanere dormienti e vitali, in condizioni ben peggiori delle piante adulte.
Secondo alcuni autori (Baskin) la dormienza può essere classificata in quattro categorie:
- d. fisiologica, es. Asteraceae, Caryophyllaceae, Brassicaceae, Plantaginaceae, Plumbaginaceae,
- Pocaceae, Scrophulariaceae;
- d. fisica, es. Convulvaceae, Fabaceae, Malvaceae;
- d. morfologica, es. Apiaceae, Amarillidaceae, Iridaceae;
- non dormienza, es. Tamarix.
Una classificazione delle alofite è basata proprio sulla germinazione e il recovery (Keiffer & Ungar, 1995). Le a. di tipo 1 sono quelle presenti sulle coste o sulle dune raramente inondate, con la pianta che tollera l’immersione solamente durante la fase di dispersione; la germinazione è tanto più probabile quanto minore è la concentrazione salina, tuttavia valori troppo elevati inibiscono completamente la possibilità di germinare. Le a. di tipo 2 sono quelle che germinano solo in condizioni di bassa concentrazione salina. Infine le a. di tipo 3 sono quelle che germinano anche in condizioni di salinità elevate e sembra che concentrazioni notevoli stimolino la germinazione stessa (Salicornia e Suaeda).
Il livello di alotolleranza del seme è variabile all’interno di una popolazione della stessa specie, quindi la fonte da cui il seme proviene sembra essere fondamentale per la determinazione del successo riproduttivo in condizioni di stress salino. Tale livello di alotolleranza risulta ancora maggiore entro specie dello stesso genere. Un esempio interessante è quello della specie Hordeum marinum, una graminacea terofita diffusa in ambienti estremamente salati, che ha adattato il proprio bioritmo ai cicli stagionali, con inimi valori di germinazione durante il periodo estivo quando sono ai massimi valori i livelli di salinità. La specie H. murinum invece mostra un discreto tasso di germinazione anche in presenza di concentrazioni notevoli di sali, questo evidenzia come anche all’interno dello stesso genere si possano incontrare differenze notevoli.
Nelle alofite la germinazione non è correlata unicamente alla salinità, altri fattori, come la temperatura sono molo importanti. A temperature maggiori l’effetto inibitorio sulla germinazione della salinità sembra essere amplificato. Inoltre il termofotoperiodo (alternanza luce/giorno e temperature), influenza la germinazione in molte specie di alofite. Nelle regioni Mediterranee molte alofite germinano in primavera, quando le temperature non sono ancora troppo elevate e le piogge tipiche del periodo tendono a dilavare e limitare le concentrazioni di sali.
Una caratteristica delle piante alotolleranti sono i semi eteroblastici. Questi presentano innumerevoli vantaggi in un ambiente ipervariabile come quello tipico delle alofite, che possiamo così riassumere:
- estensione del periodo di germinazione grazie a diversi gradi di dormienza;
- produzione di una persistente banca semi che favorisce una sicura riserva di plantule;
- adeguata risposta alle variazioni ambientali e conseguente vantaggio selettivo.
Il polimorfismo dei semi è correlato ad una effettiva differenza nella morfologia, per esempio essi possono essere grandi o piccoli, chiari o scuri ecc...e a diversi momenti nella produzione, e quindi della maturazione e germinazione dei semi stessi. La differenza di taglia è correlata per esempio alla posizione dei fiori nell’infiorescenza (Salicornia europea), agli stress ambientali (Salsola), all’induzione del fotoperiodo. In Zanichellia palustris sono presenti due tipi di frutti (acheni) morfofisiologicamente diversi sullo stesso individuo (Lombardi T., Bedini S., Onnis A., 1996).
UN MODELLO DI ALOFITA
Da tempo, come modello di studio delle glicofite si utilizza Arabidopsis, che è stato anche il primo vegetale ad essere sequenziato. La specie non è particolarmente tollerante agli stress salini, ma attraverso metodi di ingegneria genetica sono stati introdotti geni estranei per studiarne gli effetti, in tutti i campi della ricerca, compresi quelli riguardanti gli stress salini. Tuttavia per meglio capire la fisiologia e la biochimica delle alofite, ancora poco comprese, sarebbe meglio avere a disposizione una specie modello. Le caratteristiche di una specie simile riguardano sopratutto il ciclo vitale, che deve essere breve e ben conosciuto, la taglia deve essere non troppo grande, deve essere facilmente coltivabile ecc...Una ottima candidata sembra essere Thellungiella halophila, che cresce spontaneamente lungo le coste della Cina dell’est. Essa ha una morfologia simile ad Arabidopsis, un ciclo vitale simile, un genoma adiruttura simile e più corto di circa la metà. La pianta tollera elevate concentrazioni saline e si sta dimostrando ottima per le ricerche sulle piante alotolleranti. Un articolo specialistico sulle caratteristiche di questa specie è riportato sulla rivista Plant Physiology, Vol 138 (Titolo: Abiotic Strees and Plant Genome Evolution: Search for new Models, Amtmann A., Bohnert H. J., Bressan A. R. 2005,Vol 138, pp127-130, disponibile gratuitamente in PDF al sito della rivista).
LA PRESSIONE OSMOTICA
Per capire meglio la relazione che lega la pressione osmotica π alla presenza di determinate molecole, esaminiamo brevemente la relazione che lega appunto la pressione osmotica π ai soluti, secondo la legge di Vant Hòff,per soluzioni diluite:
π = n R T
dove n è il numero di moli per Kg di solvente (o molalità; nel caso di soluzioni diluite molalità e molarità coincidono), R è la costante dei gas e infine T è la temperatura assoluta. Si può osservare che π dipende solo dalla concentrazione dei soluti e non dalla natura del soluto. Quindi soluzioni con soluti diversi ma con lo stesso numero di molecole, hanno la stessa pressione osmotica. Il vantaggio sta nel fatto che se vengono idrolizzati dei polimeri, la pressione osmotica sale, all’opposto se vengono condensati dei monomeri la pressione scende.
ARTICOLI CORRELATI
- GENESI E CLASSIFICAZIONE DELLE DUNE COSTIERE
- LA FLORA DEGLI STAGNI
- LE DUNE COSTIERE DEI GRANDI LAGHI AMERICANI
- LA VEGETAZIONE DEGLI AMBIENTI DUNALI
- LE PIANTE PSAMMOFILE
- CLASSIFICAZIONE DELLE SPIAGGE
BIBLIOGRAFIA
- La classificazione riportata è tratta dal testo: Handbook of plant and crop stress. pp128.
- Jennifer Seaman, Mechanism of salt tolerance in halophytes: can crop plants resistance to salinity be improved? Disponibile on line.
- Jian-Kang-Zhu, Plant salt tolerance, Trends in Plant Science, Vol. 6 n.2, pp 66-71. 2001.
- Keiffer & Ungar, 1995
- Xiamou Miu et al. NaCl regulation of plasma membrane H+ ATPase gene expression in a Glicophyte and halophyte, Plant Physiology 103, pp 713-718. 1993
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
